 |
|
МЕНЮ
|
Контрольная работа: Механизмы реализации путей гибели клеткиВ плазмолемме эритроцита идентифицировано 15 главных белков с молекулярной массой 15-250 КД. Более 60 % всех белков составляют примембранный белок спектрин, мембранные белки - гликофорин и полоса спектрин составляет 25 % массы всех мембранных и примембранных белков эритроцита, является белком цитоскелета, связанным с цитоплазматической стороной плазмолеммы, участвует в поддержании двояковогнутой формы эритроцита Молекула спектрина имеет вид палочки длиной 100 нм, состоящей из 2 полипептидных цепей: а-спектрина B40 КД) и р-спектрина B20 КД). Концы сформированных из них тетрамеров связаны с короткими актиновыми филаментами цитоплазмы и белком полосы 4.1, образуя "узловой комплекс" (рис.13). Цитоскелетный белок полосы 4.1, связывающий спектрин и актин, одновременно соединяется с белком гликофорином. На внутренней цитоплазматической поверхности плазмолеммы образуется гибкая сетевидная структура, которая поддерживает форму эритроцита и противостоит давлению при прохождении его через тонкий капилляр (см. рис.14). Доказано, что при наследственной аномалии спектрина эритроциты имеют сферическую форму. При недостаточности спектрина в условиях анемии эритроциты также принимают сферическую форму. Соединение спектринового цитоскелета с плазмолеммой обеспечивает внутриклеточный белок анкирин. Анкирин связывает спектрин с трансмембранным белком плазмолеммы (полоса 3). Гликофорин - трансмембранный белок C0 КД), который пронизывает плазмолемму в виде одиночной спирали, и его большая часть выступает на наружной поверхности эритроцита, где к нему присоединены 15 отдельных цепей олигосахаридов, которые в сумме составляют 60 % массы гликофорина и несут отрицательные заряды.
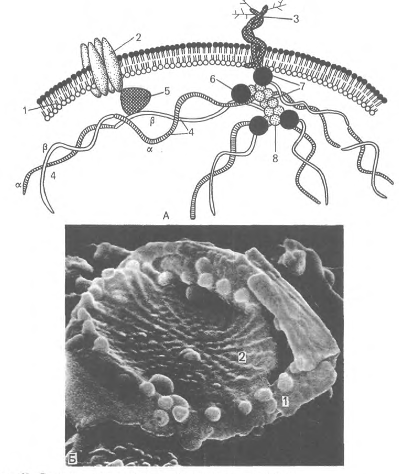
Рис.13. Цитоскелет фибробласта и эритроцит Гликофорины относятся к классу мембранных гликопротеинов, которые выполняют рецепторные функции. Гликофорины обнаружены только в эритроцитах. Полоса 3 представляет собой-трансмембранный гликопротеид A00 КД), полипептидная цепь которого много раз пересекает бислой липидов. Этот гликопротеид участвует в обмене О2 и СО2, которые связывают гемоглобин - основной белок цитоплазмы эритроцита. Эритроциты в легких отдают СО2 путем замены анионов НСО" на СГ. Белок полосы 3 обеспечивает этим анионам трансмембранный проход через гидрофильные "поры", окруженные гидрофобными липидными зонами. Таким образом формируются водные ионные каналы.
Рис.14. Строение плазмолеммы и цитоскелета эритроцита. А - схема: 1 - плазмолемма; 2 - белок полосы 3; 3 - гликофорин; 4 - сттектрин {а - и р - цепи); 5 - анкирин; 6 - белок полосы 4.1; 7 - узловой комплекс; 8 - актин; Б - плазмолемма и цитоскелет эритроцита в сканирующем электронном микроскопе. - плазмолемма: 2 - сеть спектрина. Олигосахариды гликолипидов и гликопротеидов образуют гликокаликс. Они определяют антигенный состав эритроцитов, т.е. наличие в них агглютиногенов. На поверхности эритроцитов выявлены агглютиногены А и В, в состав которых входят полисахариды, содержащие амнносахара и глюкуроновую кислоту. Они обеспечивают агглютинацию (склеивание) эритроцитов под влиянием соответствующих белков плазмы крови - а - и р-агглютининов, находящихся в составе фракции у-глобулинов. По содержанию агглютиногенов и агглютининов различают 4 группы крови: в крови А (0) группы отсутствуют агглютиногсны А и В, но имеются а - и р-агтлютинины; в крови А (П) группы имеются агглютиноген А и р-агглютинин; в крови В (Ш) группы содержатся В-агглютиноген и а-агглютинин; в крови AB (IV) группы имеются агглютиногены А и В и нет агглютининов. При переливании крови для предотвращения гемолиза (разрушение эритроцитов) нельзя допускать вливания реципиентам эритроцитов с агглютиногенами А или В, имеющим а - или р-агглютинины. Поэтому лица с (0) A группой крови являются универсальными донорами, т.е. их кровь может быть перелита всем людям с другими группами крови. Соответственно лица с AB (FV) группой крови являются универсальными реципиентами, т.е. им можно перелить любую группу крови. На поверхности эритроцитов имеется также резус-фактор (Rh-фактор) - агглютиноген. Он присутствует у 86 % людей; у 14 % отсутствует (резус-отрицательные). Переливание резус-положительной крови резус-отрицательному пациенту вызывает образование резус-антител и гемолиз эритроцитов. Агглютинация эритроцитов свойственна нормальной свежей крови, при этом образуются так называемые "монетные столбики". Это явление связано с потерей заряда плаэмолеммы эритроцитов. Скорость оседания (агглютинации) эритроцитов (СОЭ) в 1 ч у здорового человека составляет 4-8 мм у мужчин и 7-10 мм у женщин. СОЭ может значительно изменяться при заболеваниях, например при воспалительных процессах, и поэтому служит важным диагностическим признаком. В движущейся крови эритроциты отталкиваются из-за наличия на их плазмолемме одноименных отрицательных зарядов. Поверхность плазмолеммы одного эритроцита составляет около 130 мкм2 крови. При окрашивании мазка крови азур И-эозином по Романовскому - Гимзе большинство эритроцитов приобретают оранжево-розовый цвет (оксифильны), что обусловлено высоким содержанием в них гемоглобина. В небольшой части эритроцитов A-5 %), являющихся более молодыми формами, сохраняются остатки органелл (рибосомы, гранулярный, эндоплазматический ретикулум), которые проявляют базофилию. Такие эритроциты окрашиваются как кислыми красителями (эозин), так и основными (азур II) и называются полихроматофилъными. При специальной суправитальной окраске (бриллиант-крезилфиолетовым) в них выявляются зернисто-нитчатые структуры, поэтому их называют ретикулоцитами. Эритроциты различаются по степени насыщенности гемоглобином. Среди них выделяются нормохромные, гипохромные и гиперхромные, соотношение между которыми значительно изменяется при заболеваниях. Количество гемоглобина в одном эритроците называют цветным показателем. Электронно-микроскопически гемоглобин выявляется в гиалоплазме эритроцита в виде многочисленных плотных гранул диаметром 4-5 нм. Гемоглобин - это сложный белок F8 КД, состоящий из 4 полипептидных цепей глобина и гема (железосодержащий порфирин), обладающий высокой способностью связывать кислород. В норме у человека содержится два типа гемоглобина - НЬА и HbF. Эти гемоглобины различаются составом аминокислот в глобиновой (белковой) части. У взрослых людей в эритроцитах преобладает НЬА, (от англ. adult - взрослый), составляя 98 %. Он содержит две а-глобиновые цепи и две C-глобиновые цепи, включающие 574 аминокислоты. HbF, или фетальный гемоглобин (от англ. foetus - плод), составляет у взрослых около 2 % и преобладает у плодов. К моменту рождения ребенка HbF составляет около 80 %, а НЬА только 20 %. Эти гемоглобины отличаются составом аминокислот в глобиновой (белковой) части. Железо (Fe2+) в геме может присоединять О2 в легких (в таких случаях образуется оксигемоглобин - НЬО2) и отдавать его в тканях путем диссоциации НЬО, на кислород (О2) и НЬ; валентность Fe2+ не изменяется. При ряде заболеваний (гемоглобинозы, гемоглобинопатии) в эритроцитах появляются другие виды гемоглобинов, которые характеризуются изменением аминокислотного состава в белковой части гемоглобина. В настоящее время выявлено более 150 видов аномальных гемоглобинов. Например, при серповидно-клеточной анемии имеет место генетически обусловленное повреждение в C-цепи гемоглобина - глютаминовая кислота, занимающая 6-е положение в полипептидной цепи, заменена на аминокислоту валин. Такой гемоглобин обозначается как HbS (от англ. sickle - серп), так как в условиях понижения парциального давления О2 он превращается в тектоидное тело, придавая эритроциту форму серпа. В ряде стран тропического пояса определенный контингент людей являются гетерозиготными для серповидных генов, а дети двух гетерозиготных родителей по законам наследственности дают либо нормальный тип B (5%), либо бывают гетерозиготными носителями, и 25 % страдают серповидноклеточной анемией. Гемоглобин способен связывать О2 в легких, при этом образуется оксигемоглобин, который транспортируется ко всем органам и тканям и там отдает О2. В тканях выделяемая СО поступает в эритроциты и соединяется с НЬ, образуя карбоксигемоглобин. При разрушении эритроцитов (старых или при воздействии различных факторов - токсины, радиация и др.) гемоглобин выходит из клеток, и это явление называется гемолизом. Старые эритроциты разрушаются макрофагами главным образом в селезенке, а также в печени и костном мозге, при этом НЬ распадается, а высвобождающееся из железосодержащего гема железо используется для образования новых эритроцитов.
Рис 15. Электронная микрофотография гемолиза эритроцитов и образование их “теней” В макрофагах НЬ распадается на пигмент билирубин и гемосидерин - аморфные агрегаты, содержащие железо. Железо гемосидерина связывается с трансферрином - негеминовым белком плазмы, содержащим железо, и захватывается специальными макрофагами костного мозга. В процессе образования эритроцитов (эритропоэз) эти макрофаги передают трансферрин в формирующиеся эритроциты, что послужило основанием назвать их клетками-кормилками. В цитоплазме эритроцитов содержатся ферменты анаэробного гликолиза, с помощью которых синтезируются АТФ и НАДН, обеспечивающие энергией главные процессы, связанные с переносом О2 и СО2, а также поддержание осмотического давления и перенос ионов через плазмолемму эритроцита. Энергия гликолиза обеспечивает активный транспорт катионов через плазмолемму, поддержание оптимального соотношения концентрации К+ и Na+ в эритроцитах и плазме крови, сохранении формы и целостности мембраны эритроцита. НАДН участвует в метаболизме НЬ, предотвращая окисление его в метгемоглобин. Эритроциты участвуют в транспорте аминокислот и полипептидов, регулируют их концентрацию в плазме крови, т.е. выполняют роль буферной системы. Постоянство концентрации аминокислот и полипептидов в плазме крови поддерживается с помощью эритроцитов, которые адсорбируют их избыток из плазмы, а затем отдают различным тканям и органам. Таким образом, эритроциты являются подвижным депо аминокислот и полипептидов. Сорбционная способность эритроцитов связана с состоянием газового режима (парциальное давление О2 и СО2 - Ро, Рсо): в частности, при действии О2 наблюдаются выход аминокислот из эритроцитов и увеличение их содержания в плазме. 2.3.2 Тромбоциты\кровяные пластинкиМелкие дисковидные двояковыпуклые безъядерные постклеточные структуры диаметром 2-4 мкм, циркулирующие в крови. Они образуются в красном костном мозге в результате фрагментации участков цитоплазмы мегакариоцитов (гигантских клеток костного мозга), поступают в кровь, в которой находятся в течение 5-10 дней, после чего фагоцитируются макрофагами, преимущественно в селезенке и легком. Часть тромбоцитов разрушается за пределами сосудистого русла, куда они попадают при повреждении стенки сосудов. Общее количество тромбоцитов в крови взрослого человека 0,8-2,4 *1012; из этого числа около 15 % обновляются ежедневно. В норме в крови циркулирует 2/3 общего числа тромбоцитов, а 1/3 находится вне циркуляции в красной пульпе селезенки. На мазках крови тромбоциты вследствие агрегации обычно выявляются в виде скоплений. Концентрация тромбоцитов в крови равна 200-400 тыс. /мкл (200-400 *109 /л) крови. Функции тромбоцитов осуществляются как внутри сосудистого русла, так и вне его. К ним относятся: 1) Остановка кровотечения при повреждении стенки сосудов (первичный гомеостаз) - основная функция тромбоцитов; 2) Обеспечение свертывания крови - (гемокоакуляция) - вторичный гомеостаз (совместно с эндотелием кровеносных сосудов и плазмой крови); 3) Участие в реакциях заживления ран (в первую очередь, повреждений сосудистой стенки) и воспаления; 4) Обеспечение нормальной функции сосудов, в частности, их эндотелиальной выстилки (ангиотрофическая функция). Строение тромбоцитов. Тромбоцит окружен плазмолеммой и включает 1. светлую прозрачную наружную часть, называемую гиаломером, 2. центральную окрашенную часть, содержащую аэурофильные гранулы - грануломер. 3. В некоторых случаях выявляются небольшие псевдоподии, выступающие из периферической части гиаломера.
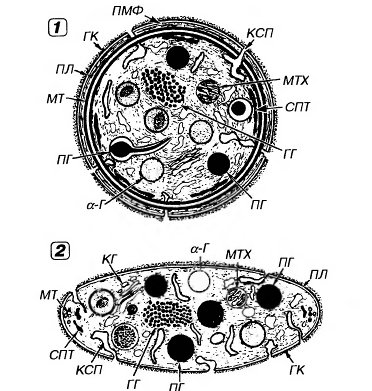
Рис.16. Ультраструктура тромбоцита.1 - сечение в экваториальной плоскости, 2 - поперечный разрез, ПЛ - плазмолемма, ГК - гликокалис, КСП - канальцы, связанные с поверхностью, СПТ - система плотных трубочек, МТ - микротрубочки, ПМФ - подмембранные микрофиламенты, ГГ - гликоген, КГ - комплекс Гольджи, МТХ - митохондрия, α-Г - α-гранулы, ПГ - плотные гранулы. 4. Плазмолемма тромбоцитов покрыта снаружи толстым (от 50 до 150-200 нм) слоем гликокалиса с высоким содержанием гликозаминогликанов и гликопротеинов (см. рисунок 17). 5. Она содержит многочисленные рецепторы, посредующие действие веществ, активирующих и ингибирующих функции тромбоцитов, обуславливающие их прикрепление (адгезию) к эндотелию сосудов и агрегацию (склеивание друг с другом). 6. Наиболее важными из них в функциональном отношении являются рецепторные гликопротеины Ib (GP Ib), IIb (GP IIb), IIIa (GP IIIa), рецепторы к АДФ, адреналину, тромбину, фактору Ха, фактору агрегации тромбоцитов (ФАТ), коллагену.
Гиаломер содержит две системы трубочек (канальцев) и большую часть элементов цитоскилета (см. рисунок 17): 1. Система канальцев, связанных с поверхностью (открытая система канальцев), представлена гладкими анастомозирующими трубочками, которые открываются в инвагинации, образованные плазмалеммой. Функция этой системы канальцев, скорее всего, связана с процессами поглощения и выведения веществ, в частности, она облегчает экзоцитоз содержимого гранул тромбоцитов. 2. Система плотных трубочек образуется комплексом Гольджи мегакариоцитов. Она представлена узкими мембранными трубочками заполненными плотным зернистым содержимым, которые располагаются непосредственно под кольцом микротрубочек. Или разбросаны по цитоплазме. Предполагают, что они накапливают и выделяют Са2+, т.е. являются аналогом саркоплазматической сети мышечных клеток. Их связывают так же с выработкой простагландинов. Цитоскелет тромбоцитов представлен микротрубочками, микрофиламентами и промежуточными филаментами. Микротрубочки в количестве 4-15 располагаются по перефирии цитоплазмы и формируют мощный пучек (краевое кольцо), служащий жестким каркасом и способствующий поддержанию формы тромбоцитов. Микрофиламенты, образованные актином, многочисленны (актин составляет 25% белка тромбоцитов), располагаются по всей цитоплазме в виде коротких нитей; в гиаломере они концентрируются между пучком микротрубочек и плазмалеммой и образует подмембранный аппарат. Последний участвует в образовании выпячиваний цитоплазмы при движении и агрегации тромбоцитов. Актиновые филаменты связаны в единую систему посредством белков α - актина, миозина и тропомиозина, а с плазмалеммой - с помощью белка филамина. Промежуточные филаменты образованы белком виментином и располагаются примущественно под плазмалеммой. Грануломер содержит митохондрии, частицы гликогена, отдельные рибосомы, единичные короткие цистерны грЭПС, элементы комплекса Гольджи и гранулы нескольких типов. α - гранулы - самые крупные (диаметр 300-500 нм), с умеренно плотным матриксом, в котором содержаться: фибриноген, фибронектин, тромбоспондин (белок сходный с актомиозином), тромбоглобулин, тромбоцитарный фактор роста (ТРФР), ЭФР, ТФРβ, фактор свертывания V и фактор Виллебанда (белок переносчик фактора VIII свертывания), а так же ряд других белков. Составляют большую часть гранул, окрашивающихся азуром. δ - гранулы (плотные гранулы или тельца) - немногочисленные (до 5) мембранные пузырьки диаметром 250-300 нм с плотным матриксом, который иногда располагается в них эксцентрично. Матрикс содержит АДФ, АТФ, Са2+, Mg2+, пирофосфат, гистамин, серотонин. Последний не синтезируется тромбоцитами, а поглощается ими из крови. γ - гранулы мелкие (диаметр 200-250 нм) пузырьки, содержащие гидролитические ферменты. Рассматриваются как лизосомы. 3. Иммунитет Под иммунитетом понимается невосприимчивость, слабовосприимчивость, сопротивляемость организма инфекциям и инвазиям чужеродных организмов (в том числе - болезнетворных микроорганизмов) и относительная устойчивость к вредным веществам. В более широком смысле это - способность организма противостоять изменению его нормального функционирования под воздействием внешних факторов. Иммунитет - это внутренняя система организма, которая обеспечивает его защиту от вредных воздействий внешней среды. Эта система защищает как основные как внешние его параметры, так и функции жизнедеятельности его органов и тканей. У большинства живых организмов существуют механизмы неспецифической резистентности, которую также называют врождённым, конституциональным или видовым иммунитетом. У челюстноротых позвоночных появляется качественно новая система приобретённого иммунитета, позволяющая реагировать на конкретный чужеродный биоматериал и запоминать его, существенно усиливая интенсивность ответной реакции на вторжение при повторном контакте с этим же материалом. Различают два типа иммунитета: специфический и неспецифический. Специфический иммунитет носит индивидуальный характер и формируется на протяжении всей жизни человека в результате контакта его иммунной системы с различными микробами и антигенами. Специфический иммунитет сохраняет память о перенесенной инфекции и препятствует ее повторному возникновению. Неспецифический иммунитет носит видоспецифический характер, то есть практически одинаков у всех представителей одного вида. Неспецифический иммунитет обеспечивает борьбу с инфекцией на ранних этапах ее развития, когда специфический иммунитет еще не сформировался. Состояние неспецифического иммунитета определяет предрасположенность человека к различным банальным инфекциям, возбудителями которых являются условно патогенные микробы. Невосприимчивость организма к инфекционным заболеваниям бывает специфической и неспецифической (неспецифическая резистентность). Специфическая невосприимчивость = иммунитет. Неспецифическая резистентность бывает обусловлена: 1. Видовой невосприимчивостью (например, невосприимчивость человека к болезням животных); 2. Бактерицидными факторами организма (фагоцитоз и воспаление, лизоцим, показатель pH, лихорадочные реакции) · Стерильный иммунитет (выздоровление сопровождается полным устранением микробов) · Нестерильный иммунитет (формируется т. н. носительство). · Иммуноглобулины типа M (IgM) (выделяются в первое время после контакта с инфекцией, обусловливают первичный иммунный ответ). · Антитела типа G (IgG) (защищает организм на протяжении длительного времени, участвует во вторичном иммунном ответе, при первичном заражении, пик их концентрации приходится на 2 неделю болезни). · Иммуноглобулины типа Е (IgE) обусловливают аллергические реакции - гиперчувствительность немедленного типа (ГНТ). · Иммуноглобулин типа А (защищает организм от проникновения инфекции через кожу и слизистые оболочки, в большом количестве содержится в грудном молоке). |


Copyright © 2012 г.
При использовании материалов - ссылка на сайт обязательна.